
 Τι είναι το εγωιστικό γονίδιο;
Τι είναι το εγωιστικό γονίδιο;
Δεν είναι απλώς ένα ξεχωριστό φυσικό κομμάτι DNA. Όπως στην αρχέγονη σούπα, είναι όλα τα αντίγραφα ενός συγκεκριμένου κομματιού DNA που υπάρχουν σε όλο τον κόσμο. Αν επιτρέπαμε στον εαυτό μας να μιλά για τα γονίδια σαν να είχαν συνειδητούς σκοπούς (να είμαστε όμως σίγουροι πως αν θέλαμε θα μεταφράζαμε αυτή τη ρευστή γλώσσα σε περισσότερο αξιόπιστους όρους), θα ρωτούσαμε : Τι «θέλει» να κάνει ένα συγκεκριμένο εγωιστικό γονίδιο ;
Η απάντηση είναι ότι προσπαθεί να εξαπλωθεί μέσα στη γονιδιακή δεξαμενή. Κι αυτό το κάνει με το να υποβοηθεί τον προγραμματισμό των σωμάτων, μέσα στα οποία βρίσκεται, για να διατηρούνται στη ζωή και να αναπαράγονται. Πρέπει όμως να τονίσουμε ότι όταν λέμε «το γονίδιο» εννοούμε όλα τα όμοιά του που υπάρχουν συγχρόνως σε πολλά άτομα. Η κεντρική ιδέα αυτού του κεφαλαίου είναι ότι ένα γονίδιο μπορεί να βοηθήσει τα αντίγραφα του εαυτού του που βρίσκονται σε άλλα σώματα. Αυτό, αν πραγματικά συμβαίνει, μοιάζει με «ατομικό» αλτρουισμό, ο οποίος όμως οφείλεται στον εγωισμό του γονιδίου.
 Ας θεωρήσουμε το γονίδιο που κάνει έναν άνθρωπο αλφικό [Αλφικός ή αλβίνος ονομάζεται το άτομο με τη γενετική ανωμαλία του αλφισμού (λευκοπάθεια), η οποία εκδηλώνεται με λευκότητα του δέρματος των τριχών και ερυθρότητα των οφθαλμών (Σ.τ.μ.)]. Στην πραγματικότητα δεν υπάρχει μόνο ένα γονίδιο που προκαλεί τον αλφισμό, όμως εγώ θα μιλώ μόνο για ένα από αυτά. Αυτό το γονίδιο είναι υπολειπόμενο, δηλαδή πρέπει να υπάρχει σε διπλή δόση για να κάνει ένα άτομο αλφικό. Αυτό συμβαίνει σε έναν στους 20.000 ανθρώπους. Ενας στους 70 έχει αυτό το γονίδιο σε απλή δόση, δεν είναι όμως αλφικός. Επειδή το αλφικό γονίδιο, όπως κάθε γονίδιο, υπάρχει σε πολλά άτομα, θεωρητικά θα υποβοηθεί την επιβίωσή του στη γονιδιακή δεξαμενή αν προγραμματίζει τα σώματα στα οποία βρίσκεται να συμπεριφέρονται αλτρουιστικά στα άλλα αλφικά σώματα, επειδή και αυτά περιέχουν το ίδιο γονίδιο. Το αλφικό γονίδιο θα ήταν πολύ «ευχαριστημένο» αν πέθαιναν μερικά σώματα που το περιέχουν, με την προϋπόθεση βέβαια ότι αυτό θα βοηθούσε να επιβιώσουν άλλα σώματα που περιέχουν το ίδιο γονίδιο. Αν το αλφικό γονίδιο μπορούσε θυσιάζοντας ένα από τα σώματα στα οποία βρίσκεται να σώσει τη ζωή 10 άλλων αλφικών σωμάτων, τότε και ο θάνατος αυτού του αλτρουιστή αντισταθμίζεται με το παραπάνω από την αύξηση του αριθμού των αλφικών γονιδίων στη γονιδιακή δεξαμενή.
Ας θεωρήσουμε το γονίδιο που κάνει έναν άνθρωπο αλφικό [Αλφικός ή αλβίνος ονομάζεται το άτομο με τη γενετική ανωμαλία του αλφισμού (λευκοπάθεια), η οποία εκδηλώνεται με λευκότητα του δέρματος των τριχών και ερυθρότητα των οφθαλμών (Σ.τ.μ.)]. Στην πραγματικότητα δεν υπάρχει μόνο ένα γονίδιο που προκαλεί τον αλφισμό, όμως εγώ θα μιλώ μόνο για ένα από αυτά. Αυτό το γονίδιο είναι υπολειπόμενο, δηλαδή πρέπει να υπάρχει σε διπλή δόση για να κάνει ένα άτομο αλφικό. Αυτό συμβαίνει σε έναν στους 20.000 ανθρώπους. Ενας στους 70 έχει αυτό το γονίδιο σε απλή δόση, δεν είναι όμως αλφικός. Επειδή το αλφικό γονίδιο, όπως κάθε γονίδιο, υπάρχει σε πολλά άτομα, θεωρητικά θα υποβοηθεί την επιβίωσή του στη γονιδιακή δεξαμενή αν προγραμματίζει τα σώματα στα οποία βρίσκεται να συμπεριφέρονται αλτρουιστικά στα άλλα αλφικά σώματα, επειδή και αυτά περιέχουν το ίδιο γονίδιο. Το αλφικό γονίδιο θα ήταν πολύ «ευχαριστημένο» αν πέθαιναν μερικά σώματα που το περιέχουν, με την προϋπόθεση βέβαια ότι αυτό θα βοηθούσε να επιβιώσουν άλλα σώματα που περιέχουν το ίδιο γονίδιο. Αν το αλφικό γονίδιο μπορούσε θυσιάζοντας ένα από τα σώματα στα οποία βρίσκεται να σώσει τη ζωή 10 άλλων αλφικών σωμάτων, τότε και ο θάνατος αυτού του αλτρουιστή αντισταθμίζεται με το παραπάνω από την αύξηση του αριθμού των αλφικών γονιδίων στη γονιδιακή δεξαμενή.
Μπορούμε λοιπόν να περιμένουμε τα αλφικά γονίδια να δείχνουν ιδιαίτερα καλή συμπεριφορά το ένα στο άλλο ; Στην πραγματικότητα η απάντηση είναι μάλλον αρνητική. Για να εξηγήσουμε αυτή την απάντηση, πρέπει να σταματήσουμε προσωρινά να μιλάμε για συνειδητά γονίδια, γιατί στο τωρινό πλαίσιο συζήτησης αυτό θα μας οδηγούσε σίγουρα σε πλάνες. Πρέπει να μιλήσουμε με όρους περισσότερο αξιόπιστους, αν και περίπλοκους. Στην πραγματικότητα, τα αλφικά γονίδια δεν «θέλουν» να επιβιώσουν ή να βοηθήσουν άλλα αλφικά γονίδια. Αν όμως κάποιο αλφικό γονίδιο τύχαινε να προκαλέσει την αλτρουιστική συμπεριφορά των σωμάτων στα οποία βρίσκεται απέναντι σε άλλα αλφικά άτομα, τότε αυτόματα, θέλοντας και μη, το αποτέλεσμα θα ήταν να εξαπλωθεί στη γονιδιακή δεξαμενή. Όμως αυτό θα συμβεί αν το γονίδιο έχει δύο αποτελέσματα ανεξάρτητα μεταξύ τους. Όχι μόνο να τους προσδίδει τη γνωστή λευκότητα αλλά και να τα παρακινεί να φέρονται αλτρουιστικά στα άτομα με έντονη ωχρότητα. Ένα τέτοιο γονίδιο «διπλού αποτελέσματος», αν υπήρχε, θα ήταν πολύ επιτυχημένο μέσα στον πληθυσμό. Όπως τόνισα στο τρίτο κεφάλαιο, στην πραγματικότητα τα γονίδια έχουν πολλαπλά αποτελέσματα. Θεωρητικά, είναι δυνατό ένα γονίδιο να δίνει στα άτομα που το περιέχουν μια ορατή ετικέτα, ας πούμε «ωχρή επιδερμίδα», «πράσινη γενειάδα» ή κάποιο άλλο ευδιάκριτο χαρακτηριστικό, αλλά και να τα παρακινεί να φέρονται ιδιαίτερα καλά στα άλλα άτομα που δείχνουν την ίδια ευδιάκριτη ετικέτα. Είναι δυνατό, αν και όχι ιδιαίτερα πιθανό.
Εξίσου πιθανό είναι η πράσινη γενειάδα να συσχετίζεται με την ανάπτυξη γαμψών νυχιών ή με κάποιο άλλο χαρακτηριστικό, και η συμπάθεια για τις πράσινες γενειάδες να συμβαδίζει με μια ανικανότητα στην αναγνώριση κάποιου αρώματος. Είναι μάλλον απίθανο ένα και το αυτό γονίδιο να δίνει ταυτόχρονα τη σωστή ετικέτα και το σωστό είδος αλτρουισμού. Εντούτοις, αποτελεί θεωρητική δυνατότητα και θα μπορούσε να ονομαστεί «Αλτρουιστικό Αποτέλεσμα της Πράσινης Γενειάδας».
 Μια κάποια ετικέτα, λόγου χάρη, η πράσινη γενειάδα, είναι απλώς ένας τρόπος με τον οποίο ένα γονίδιο μπορεί να «αναγνωρίσει» σε άλλα άτομα την ύπαρξη αντιγράφων του. Μήπως όμως υπάρχουν και άλλοι τρόποι ; Ενας ιδιαίτερα άμεσος τρόπος είναι ο εξής : Ο κάτοχος ενός αλτρουιστικού γονιδίου μπορεί να αναγνωριστεί απλούστατα από το γεγονός ότι κάνει πράξεις αλτρουισμού. Ένα γονίδιο θα ευημερούσε στη γονιδιακή δεξαμενή αν «έλεγε» στη «γλώσσα» του τα εξής : «Σώμα, αν ο Α πνίγεται ενώ προσπαθεί να σώσει κάποιον άλλον που πνίγεται, πέσε στο νερό και σώσε τον Α». Αυτή η συμπεριφορά του γονιδίου είναι σωστή επειδή η πιθανότητα να περιέχει ο Α το ίδιο αλτρουιστικό γονίδιο που σώζει ζωές είναι μεγαλύτερη από τον μέσο όρο. Το γεγονός ότι ο Α προσπαθεί να σώσει κάποιον άλλο είναι μια ετικέτα όπως η πράσινη γενειάδα. Είναι λιγότερο αυθαίρετη, εξακολουθεί όμως να μην είναι πειστική.
Μια κάποια ετικέτα, λόγου χάρη, η πράσινη γενειάδα, είναι απλώς ένας τρόπος με τον οποίο ένα γονίδιο μπορεί να «αναγνωρίσει» σε άλλα άτομα την ύπαρξη αντιγράφων του. Μήπως όμως υπάρχουν και άλλοι τρόποι ; Ενας ιδιαίτερα άμεσος τρόπος είναι ο εξής : Ο κάτοχος ενός αλτρουιστικού γονιδίου μπορεί να αναγνωριστεί απλούστατα από το γεγονός ότι κάνει πράξεις αλτρουισμού. Ένα γονίδιο θα ευημερούσε στη γονιδιακή δεξαμενή αν «έλεγε» στη «γλώσσα» του τα εξής : «Σώμα, αν ο Α πνίγεται ενώ προσπαθεί να σώσει κάποιον άλλον που πνίγεται, πέσε στο νερό και σώσε τον Α». Αυτή η συμπεριφορά του γονιδίου είναι σωστή επειδή η πιθανότητα να περιέχει ο Α το ίδιο αλτρουιστικό γονίδιο που σώζει ζωές είναι μεγαλύτερη από τον μέσο όρο. Το γεγονός ότι ο Α προσπαθεί να σώσει κάποιον άλλο είναι μια ετικέτα όπως η πράσινη γενειάδα. Είναι λιγότερο αυθαίρετη, εξακολουθεί όμως να μην είναι πειστική.
Άραγε υπάρχουν πειστικότεροι τρόποι με τους οποίους τα γονίδια θα μπορούσαν να «αναγνωρίσουν» τα αντίγραφά τους σε άλλα άτομα ;
Η απάντηση είναι καταφατική. Είναι εύκολο να δείξουμε ότι οι στενοί συγγενείς (γονείς και παιδιά) έχουν περισσότερες πιθανότητες από το μέσον όρο να έχουν κοινά γονίδια. Από πολύ καιρό έχει γίνει σαφές ότι αυτή είναι η αιτία του συχνά παρατηρούμενου αλτρουισμού των γονέων για τα παιδιά τους. Ο R.A. Fisher, o J.B.S. Haldane, και ειδικότερα ο W.D. Hamilton, κατάλαβαν ότι το ίδιο γίνεται και με άλλους στενούς συγγενείς – αδελφούς και αδελφές, ανιψιούς και ανιψιές, και πρωτοξάδελφα. Αν κάποιο άτομο θυσιαστεί για να σώσει 10 στενούς συγγενείς, χάνεται βέβαια ένα αντίγραφο του κοινού αλτρουιστικού γονιδίου, σώζεται όμως μεγαλύτερος αριθμός αντιγράφων αυτού του γονιδίου.
Η έκφραση «μεγαλύτερος αριθμός» είναι κάπως ασαφής, όπως και η έκφραση «στενοί συγγενείς». Ο Hamilton έδειξε ότι μπορούμε να κάνουμε κάτι καλύτερο. Οι δύο μελέτες που δημοσίευσε το 1964 συγκαταλέγονται στις σημαντικότερες συνεισφορές στην κοινωνική ηθολογία και αδυνατώ να καταλάβω γιατί οι ηθολόγοι δεν τους έδωσαν την απαιτούμενη προσοχή (το όνομα του Hamilton δεν αναγράφεται ούτε στα ευρετήρια των δύο μεγάλων εγχειριδίων ηθολογίας που εκδόθηκαν το 1970. Ευτυχώς, υπάρχουν πρόσφατες ενδείξεις αναζωπύρωσης του ενδιαφέροντος για τις ιδέες του). Οι εργασίες του Hamilton είναι μάλλον μαθηματικές αλλά είναι εύκολο να συλλάβουμε διαισθητικά τις βασικές αρχές της θεωρίας του χωρίς μαθηματικά, μολονότι η υπεραπλούστευση ζημιώνει. Αυτό που θέλουμε να υπολογίσουμε είναι η πιθανότητα δύο άτομα, ας πούμε δύο αδελφές, να έχουν κοινό ένα συγκεκριμένο γονίδιο. Για ευκολία, θα εξετάσουμε την περίπτωση γονιδίων που σπανίζουν σε όλη τη γονιδιακή δεξαμενή.
Οι περισσότεροι άνθρωποι, είτε έχουν κάποια συγγένεια μεταξύ τους είτε όχι, έχουν το γονίδιο «για να μην είναι αλφικοί». Ο λόγος για τον οποίο αυτό το γονίδιο είναι τόσο διαδεδομένο είναι ότι στη φύση οι αλφικοί έχουν μικρότερες πιθανότητες να επιβιώσουν απ’ ό, τι οι μη αλφικοί. Για παράδειγμα, ο Ηλιος θαμπώνει τους αλφικούς και μειώνει την ικανότητά τους να δουν ένα αρπακτικό που πλησιάζει. Εδώ δεν θα προσπαθήσουμε να εξηγήσουμε γιατί στη γονιδιακή δεξαμενή αφθονούν τα προφανώς «καλά» γονίδια, όπως το γονίδιο του μη αλφισμού.
 Σκοπός μας είναι να εξηγήσουμε την επιτυχία των γονιδίων ειδικά λόγω του αλτρουισμού τους. Μπορούμε συνεπώς να δεχτούμε ότι τέτοια γονίδια, τουλάχιστον στα πρώτα στάδια της εξελικτικής διαδικασίας, ήταν σπάνια. Τώρα, το σημαντικό σημείο είναι ότι ένα γονίδιο, ακόμη κι αν είναι σπάνιο στο σύνολο του πληθυσμού, είναι κοινό μέσα σε μια οικογένεια. Εγώ, όπως κι εσείς, έχουμε μερικά γονίδια που είναι σπάνια στον συνολικό πληθυσμό. Οι πιθανότητες όμως να έχουμε και οι δύο τα ίδια σπάνια γονίδια είναι πραγματικά πολύ μικρές. Εντούτοις, οι πιθανότητες να έχει η αδελφή μου κάποιο συγκεκριμένο σπάνιο γονίδιο που το έχω και εγώ είναι μεγάλες, όπως υπάρχει εξίσου μεγάλη πιθανότητα να έχει η αδελφή σας ένα σπάνιο γονίδιο κοινό με το δικό σας. Σ’ αυτή την περίπτωση η πιθανότητα είναι 50% κι αυτό εξηγείται εύκολα.__
Σκοπός μας είναι να εξηγήσουμε την επιτυχία των γονιδίων ειδικά λόγω του αλτρουισμού τους. Μπορούμε συνεπώς να δεχτούμε ότι τέτοια γονίδια, τουλάχιστον στα πρώτα στάδια της εξελικτικής διαδικασίας, ήταν σπάνια. Τώρα, το σημαντικό σημείο είναι ότι ένα γονίδιο, ακόμη κι αν είναι σπάνιο στο σύνολο του πληθυσμού, είναι κοινό μέσα σε μια οικογένεια. Εγώ, όπως κι εσείς, έχουμε μερικά γονίδια που είναι σπάνια στον συνολικό πληθυσμό. Οι πιθανότητες όμως να έχουμε και οι δύο τα ίδια σπάνια γονίδια είναι πραγματικά πολύ μικρές. Εντούτοις, οι πιθανότητες να έχει η αδελφή μου κάποιο συγκεκριμένο σπάνιο γονίδιο που το έχω και εγώ είναι μεγάλες, όπως υπάρχει εξίσου μεγάλη πιθανότητα να έχει η αδελφή σας ένα σπάνιο γονίδιο κοινό με το δικό σας. Σ’ αυτή την περίπτωση η πιθανότητα είναι 50% κι αυτό εξηγείται εύκολα.__
Ας υποθέσουμε ότι έχετε ένα μόνο αντίγραφο του γονιδίου Γ. Αυτό το γονίδιο το κληρονομήσατε από τον πατέρα ή τη μητέρα σας (για ευκολία, μπορούμε να αγνοήσουμε διάφορα σπάνια ενδεχόμενα όπως, λόγου χάρη, το Γ να είναι κάποια νέα μετάλλαξη ή να το είχαν και οι δύο γονείς σας ή ένας από τους γονείς σας να το είχε σε δύο αντίγραφα). Ας υποθέσουμε ότι το κληρονομήσατε από τον πατέρα σας. Συνεπώς, κάθε σωματικό κύτταρο του πατέρα σας περιείχε ένα αντίγραφο του Γ. Θυμηθείτε τώρα ότι όταν ο άντρας παράγει ένα σπερματοζωάριο, του δίνει τα μισά από τα γονίδιά του [Που υπάρχουν σε κάθε σωματικό κύτταρό του (Σ.τ.μ.)]. Υπάρχει λοιπόν μια πιθανότητα 50% το σπερματοζωάριο που «δημιούργησε» την αδελφή σας να είχε το γονίδιο Γ. Αν πάλι κληρονομήσατε το Γ από τη μητέρα σας, με την ίδια συλλογιστική βρίσκουμε ότι το Γ πρέπει να υπήρχε στα μισά ωάριά της. Για μια φορά ακόμη, οι πιθανότητες να έχει να έχει η αδελφή σας το Γ είναι 50%. Αυτό σημαίνει ότι αν είχατε 100 αδελφούς και αδελφές, περίπου οι 50 θα είχαν κάποιο σπάνιο γονίδιο που ίσως έχετε κι εσείς. Σημαίνει επίσης ότι αν έχετε 100 σπάνια γονίδια, περίπου τα 50 θα υπάρχουν στο σώμα καθενός από τους αδελφούς και τις αδελφές σας.
Μπορείτε να κάνετε ανάλογους υπολογισμούς για όποιο βαθμό συγγένειας θέλετε. Ενδιαφέρον παρουσιάζει η σχέση γονιού-παιδιού. Αν έχετε ένα αντίγραφο του γονιδίου Δ, η πιθανότητα ένα από τα παιδιά σας να έχει όμοιο αντίγραφο είναι 50%, γιατί τα μισά από τα γενετικά κύτταρά σας περιέχουν ένα Δ και κάθε παιδί σας έγινε από ένα γεννητικό κύτταρό σας. Αν έχετε ένα αντίγραφο του γονιδίου Ε, η πιθανότητα να είχε και ο πατέρας σας ένα Ε είναι πάλι 50% γιατί τα μισά γονίδιά σας τα κληρονομήσατε από τον πατέρα σας και τα υπόλοιπα από τη μητέρα σας. Για λόγους ευκολίας χρησιμοποιούμε ένα δείκτη συγγένειας (Δ.Σ.). Εκφράζει την πιθανότητα που έχει ένα γονίδιο να είναι κοινό σε δύο συγγενείς. Ο δείκτης συγγένειας δύο αδελφών είναι ½ γιατί τα μισά γονίδια του ενός αδελφού μπορεί να υπάρχουν και στον άλλο. Ο αριθμός αυτός είναι μέσος όρος : επειδή στη μειωτική διαίρεση τα χρωματοσώματα χωρίζονται στην τύχη, είναι δυνατό συγκεκριμένα ζευγάρια αδελφών να έχουν κοινά περισσότερα ή λιγότερα γονίδια. Όμως ο Δ.Σ. μεταξύ πατέρα και παιδιού είναι πάντα ½ .
 Η εκτέλεση τέτοιων υπολογισμών από την αρχή κάθε φορά που θα χρειαστεί είναι μάλλον κουραστική. Βρέθηκε λοιπόν ένας προσεγγιστικός, άμεσος κανόνας για να υπολογίζουμε το δείκτη συγγένειας μεταξύ δύο ατόμων Α και Β. Ισως τον βρείτε χρήσιμο αν χρειαστεί να συντάξετε τη διαθήκη σας ή να εξηγήσετε εμφανείς ομοιότητες ανάμεσα στα μέλη της οικογένειάς σας. Ο κανόνας ισχύει για όλες τις απλές περιπτώσεις, αλλά, όπως θα δούμε, δεν εφαρμόζεται στην αιμομιξία και σε ορισμένα έντομα. Αναζητούμε πρώτα όλους τους κοινούς προγόνους των Α και Β. Λόγου χάρη, οι κοινοί πρόγονοι δύο πρωτοξαδέλφων είναι ο κοινός παππούς και η κοινή γιαγιά τους. Όταν βρείτε έναν κοινό πρόγονο, είναι φυσικό όλοι ο πρόγονοί του να είναι και αυτοί κοινοί πρόγονοι των Α και Β. Εντούτοις, θα τους αγνοήσουμε όλους, εκτός από τους πιο κοντινούς. Μ’ αυτή την έννοια, τα πρωτοξάδελφα έχουν μόνο δύο κοινούς προγόνους. Αν ο Β είναι κατευθείαν απόγονος του Α, λόγου χάρη δισέγγονός του, τότε ο Α είναι ο κοινός πρόγονος που αναζητούμε.
Η εκτέλεση τέτοιων υπολογισμών από την αρχή κάθε φορά που θα χρειαστεί είναι μάλλον κουραστική. Βρέθηκε λοιπόν ένας προσεγγιστικός, άμεσος κανόνας για να υπολογίζουμε το δείκτη συγγένειας μεταξύ δύο ατόμων Α και Β. Ισως τον βρείτε χρήσιμο αν χρειαστεί να συντάξετε τη διαθήκη σας ή να εξηγήσετε εμφανείς ομοιότητες ανάμεσα στα μέλη της οικογένειάς σας. Ο κανόνας ισχύει για όλες τις απλές περιπτώσεις, αλλά, όπως θα δούμε, δεν εφαρμόζεται στην αιμομιξία και σε ορισμένα έντομα. Αναζητούμε πρώτα όλους τους κοινούς προγόνους των Α και Β. Λόγου χάρη, οι κοινοί πρόγονοι δύο πρωτοξαδέλφων είναι ο κοινός παππούς και η κοινή γιαγιά τους. Όταν βρείτε έναν κοινό πρόγονο, είναι φυσικό όλοι ο πρόγονοί του να είναι και αυτοί κοινοί πρόγονοι των Α και Β. Εντούτοις, θα τους αγνοήσουμε όλους, εκτός από τους πιο κοντινούς. Μ’ αυτή την έννοια, τα πρωτοξάδελφα έχουν μόνο δύο κοινούς προγόνους. Αν ο Β είναι κατευθείαν απόγονος του Α, λόγου χάρη δισέγγονός του, τότε ο Α είναι ο κοινός πρόγονος που αναζητούμε.
Εχοντας εντοπίσει τους κοινούς προγόνους (ή τον κοινό πρόγονο) των Α και Β, υπολογίζουμε τη γενεαλογική απόστασή τους ως εξής : Αρχίζοντας από τον Α ανεβαίνουμε στο οικογενειακό δέντρο ώσπου να βρούμε έναν κοινό πρόγονο, κατόπιν κατεβαίνουμε πάλι ως τον Β. Ο ολικός αριθμός των βημάτων κατά την άνοδο και τη κάθοδο στο δέντρο είναι η γενεαλογική απόσταση. Για παράδειγμα, αν ο Α είναι θείος του Β η γενεαλογική απόσταση είναι των Α και Β είναι 3. Ο κοινός πρόγονος είναι ο πατέρας του Α (ας πούμε), δηλαδή ο παππούς του Β. Αρχίζοντας από τον Α, για να βρούμε τον κοινό πρόγονο πρέπει να ανεβούμε μια γενεά. Κατόπιν, για να φτάσουμε στον Β πρέπει να κατεβούμε δύο γενεές. Συνεπώς η γενεαλογική απόσταση είναι 1+2=3. Αφού βρούμε τη γενεαλογική απόσταση των Α και Β διαμέσου ενός συγκεκριμένου κοινού προγόνου, υπολογίζουμε το ποσοστό συνεισφοράς του κοινού προγόνου στο δείκτη συγγένειάς τους. Αυτό το κάνουμε πολλαπλασιάζοντας το ½ με τον εαυτό του για κάθε βήμα της γενεαλογικής απόστασης. Αν αυτή η απόσταση είναι 3 θα έχουμε ½ x ½ x ½ , δηλαδή ( ½ ) στη τρίτη δύναμη. Αν η απόσταση γενεών διαμέσου ενός συγκεκριμένου προγόνου περιλαμβάνει ν βήματα, το ποσοστό συγγένειας που οφείλεται σ’ αυτόν τον πρόγονο είναι ( ½ ) στη νιοστή δύναμη.
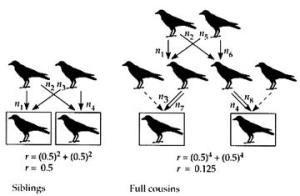 Αυτό όμως είναι μόνο ένα μέρος της συγγένειας των Α και Β. Αν έχουν περισσότερους από έναν κοινούς προγόνους, πρέπει να προσθέσουμε τον αντίστοιχο για κάθε πρόγονο αριθμό. Συνήθως η γενεαλογική απόσταση δύο ατόμων είναι η ίδια για όλους τους κοινούς προγόνους τους. Επομένως, αν βρούμε το δείκτη συγγένειας των Α και Β που οφείλεται στον έναν πρόγονο, το μόνο που μένει να κάνουμε είναι να τον πολλαπλασιάσουμε με τον αριθμό των προγόνων. Για παράδειγμα, τα πρωτοξάδελφα έχουν δύο κοινούς προγόνους και η απόσταση γενεών διαμέσου καθενός προγόνου είναι 4. Συνεπώς ο δείκτης συγγένειάς τους θα είναι : 2 x ( ½ ) στη τετάρτη δύναμη = 1/8. Αν ο Α είναι δισέγγονος του Β, η απόσταση γενεών είναι 3 και ο αριθμός των κοινών «προγόνων» είναι ένας (ο Β), τότε ο δείκτης συγγένειας είναι : 1 x ( ½ ) στη τρίτη δύναμη = 1/8. Από γενετική άποψη ένας πρωτοξάδελφός σας ισοδυναμεί με έναν δισέγγονό σας.
Αυτό όμως είναι μόνο ένα μέρος της συγγένειας των Α και Β. Αν έχουν περισσότερους από έναν κοινούς προγόνους, πρέπει να προσθέσουμε τον αντίστοιχο για κάθε πρόγονο αριθμό. Συνήθως η γενεαλογική απόσταση δύο ατόμων είναι η ίδια για όλους τους κοινούς προγόνους τους. Επομένως, αν βρούμε το δείκτη συγγένειας των Α και Β που οφείλεται στον έναν πρόγονο, το μόνο που μένει να κάνουμε είναι να τον πολλαπλασιάσουμε με τον αριθμό των προγόνων. Για παράδειγμα, τα πρωτοξάδελφα έχουν δύο κοινούς προγόνους και η απόσταση γενεών διαμέσου καθενός προγόνου είναι 4. Συνεπώς ο δείκτης συγγένειάς τους θα είναι : 2 x ( ½ ) στη τετάρτη δύναμη = 1/8. Αν ο Α είναι δισέγγονος του Β, η απόσταση γενεών είναι 3 και ο αριθμός των κοινών «προγόνων» είναι ένας (ο Β), τότε ο δείκτης συγγένειας είναι : 1 x ( ½ ) στη τρίτη δύναμη = 1/8. Από γενετική άποψη ένας πρωτοξάδελφός σας ισοδυναμεί με έναν δισέγγονό σας.
Παρομοίως, έχετε τόσες πιθανότητες να «μοιάζετε» με το θείο σας [δείκτης συγγένειας 2 x ( ½ ) στη τρίτη δύναμη = ¼ ] όσες και με τον παππού σας [Δ.Σ. 1 x ( ½ ) στο τετράγωνο = ¼ ]. Για σχέσεις τόσο μακρινές όσο ανάμεσα σε τριτοξάδελφα [2 x ( ½ ) στην ογδόη δύναμη = 1/128], πλησιάζουμε την ακραία περίπτωση ένα συγκεκριμένο γονίδιο του Α να έχει τόση πιθανότητα να βρίσκεται και στον Β όση και σε ένα τυχαίο άτομο του πληθυσμού.
Για ένα αλτρουιστικό γονίδιο, το τριτοξάδελφο δεν απέχει πολύ από οποιοδήποτε φίλο μας. Λίγο πλεονεκτεί ο δευτεροξάδελφος (Δ.Σ. = 1/32) και περισσότερο ο πρωτοξάδελφος (Δ.Σ. = 1/8). Ειδική περίπτωση είναι οι αδελφοί και οι αδελφές, και οι γονείς με τα παιδιά τους (Δ.Σ. = ½ ) και μοναδική περίπτωση οι αυθεντικοί δίδυμοι (Δ.Σ. = 1). Θείοι και θείες, ανιψιοί και ανιψιές, παππούδες και εγγόνια, και ετεροθαλείς αδελφοί και αδελφές έχουν ενδιάμεσες σχέσεις με δείκτη συγγένειας ¼ .
 Τώρα μπορούμε να μιλήσουμε ακριβέστερα για τον αλτρουισμό μεταξύ συγγενών. Ένα γονίδιο που προκαλεί την αυτοθυσία ενός ατόμου για να σώσει 5 ξαδέλφια του, δεν γίνεται πολυαριθμότερο στον πληθυσμό, πράγμα που συμβαίνει αν σώσει 5 αδέλφια του ή 10 πρωτοξάδελφα. Η ελάχιστη απαίτηση για να είναι επιτυχημένο ένα αυτοθυσιαζόμενο αλτρουιστικό γονίδιο είναι να σώσει περισσότερα από δύο αμφιθαλή αδέλφια [Αδέλφια από τον ίδιο πατέρα και την ίδια μητέρα (Σ.τ.μ.)] ή (παιδιά ή γονείς), ή περισσότερα από 4 ετεροθαλή αδέλφια (ή θείους, θείες, ανιψιούς, ανιψιές, παππούδες, εγγόνια), ή περισσότερα από 8 πρωτοξάδελφα κλπ. Κατά μέσο όρο, ένα τέτοιο γονίδιο τείνει να συνεχίζει να ζει στα σώματα των ατόμων που έσωσε ο αλτρουιστής, εφόσον είναι αρκετά ώστε να αντισταθμιστεί η απώλεια από το θάνατό του. Αν κάποιος ήταν σίγουρος ότι ένα συγκεκριμένο άτομο είναι αυθεντικός δίδυμος αδελφός του, θα έπρεπε να νοιάζεται γι’ αυτόν όσο ακριβώς και για τον εαυτό του. Κάθε γονίδιο για τον αλτρουισμό μεταξύ διδύμων υπάρχει και στους δύο, οπότε αν ο ένας θυσιαστεί για να σώσει τον άλλο, το γονίδιο συνεχίζει να ζει.
Τώρα μπορούμε να μιλήσουμε ακριβέστερα για τον αλτρουισμό μεταξύ συγγενών. Ένα γονίδιο που προκαλεί την αυτοθυσία ενός ατόμου για να σώσει 5 ξαδέλφια του, δεν γίνεται πολυαριθμότερο στον πληθυσμό, πράγμα που συμβαίνει αν σώσει 5 αδέλφια του ή 10 πρωτοξάδελφα. Η ελάχιστη απαίτηση για να είναι επιτυχημένο ένα αυτοθυσιαζόμενο αλτρουιστικό γονίδιο είναι να σώσει περισσότερα από δύο αμφιθαλή αδέλφια [Αδέλφια από τον ίδιο πατέρα και την ίδια μητέρα (Σ.τ.μ.)] ή (παιδιά ή γονείς), ή περισσότερα από 4 ετεροθαλή αδέλφια (ή θείους, θείες, ανιψιούς, ανιψιές, παππούδες, εγγόνια), ή περισσότερα από 8 πρωτοξάδελφα κλπ. Κατά μέσο όρο, ένα τέτοιο γονίδιο τείνει να συνεχίζει να ζει στα σώματα των ατόμων που έσωσε ο αλτρουιστής, εφόσον είναι αρκετά ώστε να αντισταθμιστεί η απώλεια από το θάνατό του. Αν κάποιος ήταν σίγουρος ότι ένα συγκεκριμένο άτομο είναι αυθεντικός δίδυμος αδελφός του, θα έπρεπε να νοιάζεται γι’ αυτόν όσο ακριβώς και για τον εαυτό του. Κάθε γονίδιο για τον αλτρουισμό μεταξύ διδύμων υπάρχει και στους δύο, οπότε αν ο ένας θυσιαστεί για να σώσει τον άλλο, το γονίδιο συνεχίζει να ζει.
Οι θηλυκοί αρμαδίλλοι [Μικρά θηλαστικά ζώα που ζουν στη Ν. Αμερική, το σώμα τους καλύπτεται με φολίδες και οστέϊνες πλάκες. Ανήκουν στην οικογένεια των δασυποδιδών (Σ.τ.μ.)] δίνουν αυθεντικά τετράδυμα σε κάθε γέννα τους. Απ’ όσο ξέρω, δεν έχει αναφερθεί κανένα περιστατικό ηρωικής αυτοθυσίας μεταξύ νεαρών αρμαδίλλων ενώ έχει τονιστεί ότι πρέπει να αναμένεται ισχυρός αλτρουισμός και θα άξιζε να πάει κανείς στη Λατινική Αμερική για να το διαπιστώσει.
Τώρα μπορούμε να δούμε ότι η γονική φροντίδα είναι ειδική περίπτωση αλτρουισμού μεταξύ συγγενών. Από γενετική άποψη, κάθε ενήλικο άτομο πρέπει να δείχνει τόσο ενδιαφέρον και φροντίδα για το ορφανό αδελφάκι του όσο και για το δικό του παιδί. Ο δείκτης συγγένειάς του με τα δύο παιδιά είναι ο ίδιος : ½ . Με όρους γονιδιακής επιλογής, ένα γονίδιο για αλτρουιστική συμπεριφορά της μεγαλύτερης αδελφής προς τα αδέλφια της έχει την ίδια πιθανότητα να εξαπλωθεί στον πληθυσμό με ένα γονίδιο για αλτρουισμό των γονέων προς τα παιδιά τους. Αυτό όμως, για διάφορους λόγους που θα δούμε αργότερα, είναι υπεραπλούστευση της πραγματικότητας.
Στη φύση η φροντίδα ανάμεσα σε αδελφούς και αδελφές υστερεί πολύ της φροντίδας των γονιών προς τα παιδιά τους. Αυτό που θέλω να τονίσω εδώ είναι ότι από γενετική άποψη η σχέση γονιού-παιδιού δεν έχει τίποτε το ιδιαίτερο σε σύγκριση με τη σχέση αδελφού- αδελφής. Το ότι οι γονείς κληρονομούν τα γονίδια στα παιδιά τους εδώ είναι αδύνατο μια αδελφή να δώσει γονίδια στην αδελφή της, δεν έχει σημασία, γιατί και οι δύο αδελφές παίρνουν από τους ίδιους γονείς όμοια αντίγραφα των ίδιων γονιδίων.
Μερικοί χρησιμοποιούν τον όρο επιλογή συγγενών για να ξεχωρίσουν αυτό το είδος της φυσικής επιλογής από την επιλογή ομάδων (την κατ’ επιλογή επιβίωση ομάδων) και την επιλογή ατόμων (την κατ’ επιλογή επιβίωση ατόμων). Η επιλογή συγγενών εξηγεί τον αλτρουισμό μέσα στην οικογένεια . Οσο στενότερη είναι η συγγένεια τόσο ισχυρότερη η επιλογή. Μολονότι δεν υπάρχει κανένα λάθος στον όρο «επιλογή συγγενών», δυστυχώς πρέπει να τον εγκαταλείψουμε γιατί τελευταία έχει γίνει τόσο μεγάλη κατάχρηση του όρου ώστε είναι πιθανό να προκαλέσει σύγχυση στους βιολόγους των επόμενων χρόνων.
Ο E. Wilson στο κατά τα άλλα θαυμάσιο βιβλίο του Κοινωνιοβιολογία : Η νέα σύνθεση, ορίζει την επιλογή συγγενών ως ειδική περίπτωση της επιλογής ομάδων. Δίνει ένα διάγραμμα που δείχνει καθαρά ότι τη θεωρεί ενδιάμεση περίπτωση μεταξύ της «επιλογής ατόμων» και της «επιλογής ομάδων» με τη συμβατική έννοια – την έννοια που χρησιμοποίησα στο πρώτο κεφάλαιο. Σήμερα, επιλογή ομάδων – ακόμη και με τον ορισμό του Wilson – σημαίνει επιλεκτική επιβίωση ομάδων ατόμων. Βέβαια, κατά κάποια έννοια η οικογένεια είναι ειδικός τύπος ομάδας. Όμως η βασική σκέψη του Hamilton είναι ότι η διάκριση ανάμεσα στην οικογένεια και τη μη οικογένεια δεν είναι αυστηρή αλλά ζήτημα μαθηματικής πιθανότητας. Στη θεωρία του Hamilton δεν αναφέρεται πουθενά ότι τα ζώα πρέπει να συμπεριφέρονται αλτρουιστικά σε όλα «τα μέλη της οικογένειας» και εγωιστικά σε όλους τους άλλους. Δεν υπάρχει σαφής διαχωριστική γραμμή ανάμεσα στην οικογένεια και τη μη οικογένεια. Δεν υπάρχει τρόπος να αποφανθούμε, λόγου χάρη, αν τα δευτεροξάδελφα εντάσσονται στην οικογένεια ή όχι : απλώς περιμένουμε ότι τα δευτεροξάδελφα τείνουν να δεχτούν 1/16 αλτρουισμού απ’ όσο δέχονται και τα παιδιά ή τα αμφιθαλή αδέλφια. Είναι σίγουρο πως η επιλογή συγγενών δεν είναι ειδική περίπτωση επιλογής ομάδων αλλά ειδική συνέπεια επιλογής γονιδίων.
Όμως στον ορισμό της επιλογής συγγενών που δίνει ο Wilson υπάρχει ένα σοβαρότερο μειονέκτημα. Αποκλείονται εσκεμμένα τα παιδιά : δεν υπολογίζονται ως συγγενείς ! Φυσικά, ο Wilson ξέρει πολύ καλά ότι τα παιδιά έχουν συγγένεια με τους γονείς τους. Για να εξηγήσει όμως την αλτρουιστική φροντίδα των γονέων για τα παιδιά τους, προτιμά να μην επικαλεστεί την επιλογή συγγενών. Εχει βέβαια το δικαίωμα να δίνει στις λέξεις την έννοια που του αρέσει, αλλά ο ορισμός του δημιουργεί συγχύσεις και ελπίζω ότι θα τον αλλάξει στις μελλοντικές εκδόσεις του πραγματικά σημαντικού βιβλίου του. Από γενετική άποψη, η φροντίδα των γονιών και ο αλτρουισμός αδελφού/αδελφής εξελίσσονται ακριβώς για τον ίδιο λόγο : και στις δύο περιπτώσεις υπάρχει μεγάλη πιθανότητα το αλτρουιστικό γονίδιο να βρίσκεται και στο σώμα του ευεργετούμενου.
Ζητώ την επιείκεια του μέσου αναγνώστη γι’ αυτή την οξεία κριτική και επιστρέφω γρήγορα στο κύριο θέμα. Ως τώρα έχω υπεραπλουστεύσει τα πράγματα αλλά ήρθε η ώρα να εισαγάγω μερικές τροποποιήσεις. Μίλησα από την άποψη θεμελιωδών στοιχείων, για γονίδια που θυσιάζονται για να σώσουν συγκεκριμένο αριθμό συγγενών με γνωστό δείκτη συγγένειας. Στην πραγματική ζωή είναι προφανές ότι τα ζώα δεν μετρούν πόσους συγγενείς σώζουν ούτε κάνουν νοερά τους υπολογισμούς του Hamilton, ακόμη κι αν είχαν κάποιο τρόπο να ξέρουν ποιοι είναι αδέλφια και ποιοι ξαδέλφια τους. Στην πραγματική ζωή η σίγουρη αυτοθυσία ή η σίγουρη «διάσωση» ζώων πρέπει να αντικατασταθεί από στατιστικούς κινδύνους θανάτου ή διάσωσης. Ακόμη κι ένας τριτοξάδελφός σας αξίζει να σωθεί εφόσον ο κίνδυνος που διατρέχετε για να τον σώσετε είναι πολύ μικρός. Εξάλλου, και εσείς και ο συγγενής σας που σκέφτεστε να σώσετε, σίγουρα θα πεθάνετε κάποια μέρα. Κάθε άτομο έχει μια «προσδοκία ζωής» που οποιοσδήποτε ασφαλιστής θα μπορούσε να την υπολογίσει με κάποια πιθανότητα σφάλματος. Το να σώσει κανείς τη ζωή ενός συγγενή που σε λίγο θα πεθάνει από γεράματα, έχει μικρότερη επίπτωση για το μέλλον στη γονιδιακή δεξαμενή από το να σώσει τη ζωή ενός νεαρού επίσης στενού συγγενή που έχει μπροστά του ολόκληρη ζωή.
Πρέπει να τροποποιήσουμε τους απλούς συμμετρικούς υπολογισμούς του δείκτη συγγένειας και να λογαριάσουμε τις περίπλοκες εκτιμήσεις των ασφαλιστών. Από γενετική άποψη, οι παππούδες και τα εγγόνια έχουν τους ίδιους λόγους να συμπεριφέρονται αλτρουιστικά μεταξύ τους γιατί έχουν κοινό το ¼ των γονιδίων τους. Όμως επειδή το εγγόνι έχει μεγαλύτερη προσδοκία ζωής, τα γονίδια για τον αλτρουισμό παππού προς εγγονό έχουν μεγαλύτερο επιλεκτικό πλεονέκτημα από τα αντίστοιχα γονίδια εγγονιού προς παππού. Είναι δυνατόν το καθαρό κέρδος από τη βοήθεια που παρέχεται σε κάποιο νεαρό μακρινό συγγενή να είναι μεγαλύτερο από το αντίστοιχο όφελος όταν βοηθιέται ένας ηλικιωμένος στενός συγγενής. (Φυσικά, δεν είναι υποχρεωτικό οι παππούδες να έχουν μικρότερη προσδοκία ζωής από τα εγγόνια τους. Σε είδη με μεγάλο δείκτη νηπιακής θνησιμότητας μπορεί να συμβεί το αντίθετο).
Επεκτείνοντας την αναφορά στα ασφαλιστικά μπορούμε να θεωρήσουμε ότι τα άτομα είναι ασφαλιστές που κάνουν ασφάλειες ζωής. Ένα άτομο μπορεί να επενδύσει ή να διακινδυνεύσει ένα μέρος της περιουσίας του πάνω στη ζωή ενός άλλου. Λογαριάζει τη συγγένεια που έχει μαζί του και το μέγεθος των κινδύνων που διατρέχει το συγγενικό άτομο, συγκρίνοντας την προσδοκία ζωής του ασφαλιζόμενου με τη δική του. Ακριβέστερα, αντί για «προσδοκία ζωής» θα ήταν καλύτερα να χρησιμοποιήσουμε τον όρο «προσδοκία αναπαραγωγής» ή, ακόμα καλύτερα, «γενετική ικανότητα να ωφελήσει τα γονίδιά του στο αναμενόμενο μέλλον». Τότε, για να εξελιχθεί η αλτρουιστική συμπεριφορά πρέπει ο καθαρός κίνδυνος του αλτρουιστή να είναι μικρότερος από το καθαρό όφελος του αποδέκτη πολλαπλασιαζόμενο επί τον δείκτη συγγένειας. Οι κίνδυνοι και τα οφέλη πρέπει να υπολογιστούν μ’ αυτή την περίπλοκη μέθοδο που χρησιμοποιούν οι ασφαλιστές.
Αλήθεια, τι σύνθετους υπολογισμούς είναι υποχρεωμένη να κάνει η καημένη μηχανή επιβίωσης, και ειδικότερα όταν βιάζεται ! Ο μεγάλος μαθηματικός βιολόγος J.B.S. Haldane (με μια εργασία του το 1955 πρόλαβε τον Hamilton, δεχόμενος ότι ένα γονίδιο που διασώζει στενούς συγγενείς από πνιγμό εξαπλώνεται στη γονιδιακή δεξαμενή), παρατήρησε ότι : «… Στις δύο περιπτώσεις που έσωσα κάποιους από ενδεχόμενο πνιγμό (με ελάχιστο δικό μου κίνδυνο) δεν είχα το χρόνο να κάνω τέτοιους υπολογισμούς». Ευτυχώς όμως, κι αυτό το ήξερε καλά ο Haldane, δεν χρειάζεται να παραδεχτούμε ότι οι μηχανές επιβίωσης κάνουν συνειδητά τέτοιους υπολογισμούς. Όπως χρησιμοποιούμε τον λογαριθμικό κανόνα χωρίς να συνειδητοποιούμε ότι ουσιαστικά χειριζόμαστε λογαρίθμους, έτσι το ζώο μπορεί να είναι από πριν προγραμματισμένο με τέτοιο τρόπο ώστε να συμπεριφέρεται σαν να είχε κάνει έναν πολύπλοκο υπολογισμό.
Αυτό μπορούμε εύκολα να το καταλάβουμε με το ακόλουθο παράδειγμα. Κάποιος ρίχνει ψηλά στον αέρα μια μπάλα. Αν την πιάσει λίγο αργότερα πριν πέσει στο έδαφος, είναι σαν να είχε επιλύσει ένα σύστημα διαφορικών εξισώσεων οι οποίες περιγράφουν την τροχιά της μπάλας. Μπορεί να μην ξέρει ή να μη τον ενδιαφέρει τι είναι μια διαφορική εξίσωση, αυτό όμως δεν επηρεάζει την επιδεξιότητά του να παίζει με την μπάλα. Σε κάποιο υποσυνείδητο επίπεδο συμβαίνει κάτι που λειτουργικά ισοδυναμεί με μαθηματικούς υπολογισμούς. Παρομοίως, όταν κάποιος παίρνει μια δύσκολη απόφαση αφού προηγουμένως σταθμίσει τα «υπέρ και τα κατά» και όλες τις πιθανές συνέπειες, στην πραγματικότητα κάνει κάτι που λειτουργικά ισοδυναμεί με τους πολύπλοκους υπολογισμούς των ασφαλιστών, τους οποίους μόνον ένας ηλεκτρονικός υπολογιστής θα μπορούσε να εκτελέσει.
Αν θέλαμε να προγραμματίσουμε έναν ηλεκτρονικό υπολογιστή για να μας δώσει με προσομοίωση ένα μοντέλο της μηχανής επιβίωσης που αποφασίζει αν θα συμπεριφερθεί αλτρουιστικά ή όχι, σε γενικές γραμμές θα ακολουθούσαμε την εξής διαδικασία : θα φτιάχναμε έναν κατάλογο όλων των εναλλακτικών κινήσεων που θα ήταν δυνατό να κάνει το ζώο. Κατόπιν θα προγραμματίζαμε τον πολύπλοκο υπολογισμό των ασφαλιστών για κάθε εναλλακτική συμπεριφορά. Όλα τα οφέλη θα πάρουν θετικό πρόσημο και όλοι οι κίνδυνοι αρνητικό. Πριν όμως προστεθούν οι αριθμοί που παριστάνουν τα οφέλη και τους κινδύνους, θα διορθωθούν πολλαπλασιαζόμενοι με τον κατάλληλο δείκτη συγγένειας. Χάριν απλότητας μπορούμε να αγνοήσουμε άλλους παράγοντες, λόγου χάρη όσους σχετίζονται με την ηλικία ή την κατάσταση υγείας. Επειδή ο δείκτης συγγένειας ενός ατόμου με τον εαυτό του είναι 1 (προφανώς έχει 100% των γονιδίων του), στον υπολογισμό οι κίνδυνοι και τα οφέλη ως προς τον εαυτό του δεν θα τροποποιηθούν αλλά θα ληφθούν με τις κανονικές τιμές τους. Το ολικό άθροισμα για κάθε εναλλακτικό τρόπο συμπεριφοράς θα παρουσιάζεται ως εξής:
Καθαρό όφελος τρόπου συμπεριφοράς = Προσωπικό όφελος – προσωπικός κίνδυνος + ½ του οφέλους του αδελφού – ½ του κινδύνου του αδελφού + ½ του οφέλους του άλλου αδελφού + 1/8 του οφέλους του πρωτοξάδελφου – 1/8 του κινδύνου του πρωτοξάδελφου + ½ του οφέλους του παιδιού – ½ του κινδύνου του παιδιού + κλπ.
Το αποτέλεσμα της άθροισης θα είναι ένας αριθμός που ονομάζεται τελικό καθαρό όφελος του συγκεκριμένου τρόπου συμπεριφοράς. Κατόπιν το μοντέλο του ζώου υπολογίζει το αντίστοιχο άθροισμα για όλους τους εναλλακτικούς τρόπους συμπεριφοράς του καταλόγου μας. Τελικά επιλέγει και ακολουθεί τον τρόπο συμπεριφοράς με το μεγαλύτερο καθαρό όφελος. Ακόμη κι αν όλα τα αθροίσματα βγουν αρνητικά, θα επιλέξει τη συμπεριφορά που αντιστοιχεί στο υψηλότερο άθροισμα, δηλαδή τη λιγότερο «κακή». Θυμηθείτε ότι κάθε θετική δράση συνεπάγεται κατανάλωση ενέργειας και χρόνου, που και τα δύο θα μπορούσε να χρησιμοποιηθούν για κάτι άλλο. Αν η συμπεριφορά με το μεγαλύτερο συνολικό καθαρό όφελος που θα προκύψει είναι «μην κάνεις τίποτε», το μοντέλο του ζώου δεν θα κάνει τίποτε.
Σ’ αυτό το σημείο θα δώσω ένα υπεραπλουστευμένο παράδειγμα, που αυτή τη φορά δεν είναι προσομοίωση ηλεκτρονικού υπολογιστή αλλά έχει τη μορφή υποκειμένου μονολόγου. Ας πούμε ότι είμαι ένα ζώο και βρίσκω 8 μανιτάρια. Αφού υπολογίσω τη θρεπτική αξία τους και αφαιρέσω κάτι εξαιτίας της μικρής πιθανότητας να είναι δηλητηριώδη, βγάζω το συμπέρασμα ότι κάθε μανιτάρι αξίζει +6 μονάδες (οι μονάδες είναι αυθαίρετες, όπως και τα κέρδη που αναφέραμε στο προηγούμενο κεφάλαιο). Τα μανιτάρια είναι τόσο μεγάλα ώστε δεν θα μπορούσα να φάω περισσότερα από τρία. Θα έπρεπε να ανακοινώσω την ανακάλυψή μου και να φωνάξω κι άλλους ότι «υπάρχει τροφή» ; Ποιος θα ερχόταν ; Ας πούμε ότι ήρθαν ένας αδελφός μου (Δ.Σ. ως προς εμένα ½ ), ένας ξάδελφός μου (Δ.Σ. 1/8) και ο Χ (ο δείκτης συγγένειάς του ως προς εμένα είναι τόσο μικρός ώστε πρακτικά μπορεί να θεωρηθεί ίσος με το μηδέν). Αν αποκρύψω την ανακάλυψή μου και φάω τα 3 μανιτάρια, το ολικό καθαρό όφελός μου την περίπτωση που θα έχω θα είναι 3×6=18. Για να βρω το ολικό καθαρό όφελός μου την περίπτωση που θα κάνω την «πρόσκληση για τροφή» χρειάζεται κάποιος υπολογισμός. Τα 8 μανιτάρια θα μοιραστούν στα 4. Τρώγοντας το μερίδιό μου, δηλαδή δύο μανιτάρια, θα έχω όφελος 2×6=12 μονάδες. Όμως επειδή έχω κοινά γονίδια με τον αδελφό μου και τον ξάδελφό μου, αν φάνε κι αυτοί από δύο μανιτάρια θα έχω συνολικό όφελος (1×12)+( ½ x12)+(1/8×12)+(0x12)=19 ½ . Η αντίστοιχη τιμή για την εγωιστική συμπεριφορά ήταν 18. Η διαφορά είναι πολύ μικρή αλλά το συμπέρασμα σαφές. Πρέπει να τους φωνάξω. Σ’ αυτή τη περίπτωση ο αλτρουισμός μου θα εξυπηρετούσε τα εγωιστικά μου γονίδια.
Στο παραπάνω παράδειγμα έκανα την απλουστευμένη παραδοχή ότι το μεμονωμένο ζώο ψάχνει να βρει αυτό που εξυπηρετεί καλύτερα τα γονίδιά του. Αυτό που πραγματικά συμβαίνει είναι ότι η γονιδιακή δεξαμενή γεμίζει με γονίδια, τα οποία επηρεάζουν τα σώματα κατά τέτοιον τρόπο ώστε να συμπεριφέρονται σαν να έκαναν τους υπολογισμούς που περιγράψαμε. Όπως και να ‘χουν τα πράγματα, ο υπολογισμός συνιστά μια προκαταρκτική πρώτη προσέγγιση όσων θεωρητικά έπρεπε να συμβαίνουν. Παραλείπει πολλά στοιχεία, λόγου χάρη τις ηλικίες των ατόμων για τα οποία μιλάμε. Επίσης, αν προηγουμένως είχα γευματίσει και μου έφτανε ένα μανιτάρι, το καθαρό όφελος από τη γνωστοποίηση της ανακάλυψής μου θα ήταν μεγαλύτερο απ’ όσο αν πεινούσα. Είναι ατέλειωτες οι συνεχείς βελτιώσεις αυτού του υπολογισμού ο οποίος θα ολοκληρωνόταν σ’ έναν ιδανικό κόσμο. Όμως η αληθινή ζωή δεν βιώνεται σε ιδανικό κόσμο. Δεν μπορούμε να περιμένουμε ότι ένα αληθινό ζώο θα υπολογίσει και την τελευταία λεπτομέρεια για να καταλήξει στην άριστη απόφαση. Πρέπει λοιπόν να ανακαλύψουμε με την παρατήρηση και το πείραμα από πρώτο χέρι πως καταφέρνουν τα ζώα και κάνουν σωστές αναλύσεις κόστους και κερδών.
Για να βεβαιωθούμε πως δεν ξεστρατίσαμε πολύ χρησιμοποιώντας υποκειμενικά παραδείγματα, ας επιστρέψουμε για λίγο στη γλώσσα των γονιδίων. Τα ζωντανά σώματα είναι μηχανές επιβίωσης προγραμματισμένες από γονίδια που έχουν επιβιώσει. Τα γονίδια που επιβίωσαν ζούσαν σε συνθήκες που έτειναν κατά μέσον όρο να χαρακτηρίζουν το περιβάλλον των ειδών στο παρελθόν. Κατά συνέπεια οι «εκτιμήσεις» κόστους και κερδών βασίζονται στην «εμπειρία» παρελθόντος, όπως ακριβώς και οι αποφάσεις που παίρνει ο άνθρωπος. Όμως, σ’ αυτή την περίπτωση, η εμπειρία έχει την ειδική έννοια της γονιδιακής εμπειρίας ή, ακριβέστερα, των συνθηκών του παρελθόντος με τις οποίες ζούσαν τα γονίδια. (Επειδή τα γονίδια προικίζουν επιπροσθέτως τις μηχανές επιβίωσης με ικανότητα του «μανθάνειν» θα μπορούσαμε να πούμε ότι γίνονται κάποιες εκτιμήσεις κόστους-κερδών με βάση και την εμπειρία του ατόμου). Εφόσον οι συνθήκες δεν αλλάζουν δραστικά, οι εκτιμήσεις θα είναι ακριβείς και οι μηχανές επιβίωσης θα τείνουν να παίρνουν κατά μέσον όρο σωστές αποφάσεις. Αν όμως οι συνθήκες μεταβληθούν δραστικά, οι μηχανές επιβίωσης θα τείνουν να παίρνουν λαθεμένες αποφάσεις και τα γονίδια θα πληρώσουν τα λάθη. Το ίδιο συμβαίνει και στον άνθρωπο : όσες αποφάσεις του βασίζονται σε μη πρόσφατες πληροφορίες τείνουν να είναι λαθεμένες.
Αλλά και οι εκτιμήσεις του δείκτη συγγένειας υπόκεινται σε σφάλματα και αβεβαιότητες. Στους υπεραπλουστευμένους υπολογισμούς που κάναμε ως τώρα, μιλούσαμε σαν να ήξεραν οι μηχανές επιβίωσης ποιοι είναι οι συγγενείς τους και να γνώριζαν το βαθμό συγγένειάς τους. Στην αληθινή ζωή μια τέτοια ακριβής γνώση είναι δυνατή μόνο περιστασιακά, όμως και πάλι ο δείκτης συγγένειας υπολογίζεται ως αριθμητικός μέσος όρος. Λόγου χάρη, υποθέστε ότι ο Α και ο Β έχουν ίσες πιθανότητες να είναι ετεροθαλείς ή αμφιθαλείς αδελφοί. Οι αντίστοιχοι δείκτες συγγένειας είναι ¼ και ½ , επειδή όμως δεν ξέρουμε ακριβώς αν είναι ετεροθαλείς ή αμφιθαλείς, ο αριθμός που πρέπει να πάρουμε είναι ο μέσος όρος, δηλαδή 3/8. Αν είναι σίγουρο ότι έχουν την ίδια μητέρα αλλά η πιθανότητα να έχουν τον ίδιο πατέρα είναι 1 προς 10, τότε είναι κατά 90% σίγουρο πως είναι ετεροθαλείς αδελφοί και κατά 10% αμφιθαλείς οπότε ο πραγματικός δείκτης συγγένειας θα είναι (1/10 x ½ )+(9/10 x ¼ )= 0,275.
Όμως για ποιον είναι σίγουρο όταν λέμε «είναι κατά 90% σίγουρο» ; Εννοούμε ότι ο φυσιοδίφης ύστερα από μακρόχρονες επιτόπιες μελέτες είναι σίγουρος 90%, ή ότι τα ζώα είναι σίγουρα 90%; Με λίγη τύχη τα δύο συμπεράσματα μπορεί να είναι σχεδόν το ίδιο πράγμα. Για να το διαπιστώσουμε πρέπει πρώτα να εξετάσουμε με ποιο τρόπο θα ήταν δυνατό τα ζώα να διακρίνουν τους στενούς συγγενείς τους.
Εμείς οι άνθρωποι ξέρουμε ποιοι είναι οι συγγενείς μας επειδή μας το είπαν, επειδή δίνουμε ονόματα στις συγγένειες, επειδή κάνουμε επίσημους γάμους, επειδή θυμόμαστε και επειδή υπάρχουν τα σχετικά έγγραφα. Πολλοί ανθρωποκοινωνιολόγοι ασχολούνται με «συγγένειες» στις κοινωνίες που μελετούν. Δεν εννοούν πραγματικές γενετικές συγγένειες αλλά υποκειμενικές και πολιτιστικές. Τα ήθη των ανθρώπων και οι τελετουργίες των διαφόρων φυλών συχνά δίνουν μεγάλη έμφαση στη συγγένεια. Η λατρεία των προγόνων είναι ευρύτατα διαδεδομένη, οι οικογενειακές υποχρεώσεις και η αφοσίωση στην οικογένεια κυριαρχούν στη ζωή μας. Οι βεντέτες και οι ενδοφατριακοί πόλεμοι ερμηνεύονται εύκολα με τη γενετική θεωρία του Hamilton. Το «ταμπού» της αιμομιξίας μαρτυρεί το μεγάλο σεβασμό που τρέφουν οι άνθρωποι προς τη συγγένεια, μολονότι το γενετικό πλεονέκτημα από το ταμπού της αιμομιξίας δεν έχει καμιά σχέση με τον αλτρουισμό. Πιθανώς σχετίζεται με τα επιβλαβή αποτελέσματα των υπολειπόμενων γονιδίων τα οποία εμφανίζονται κατά την ενδογαμία. (Για κάποιο λόγο, σε πολλούς ανθρωπολόγους δεν αρέσει αυτή η εξήγηση). Πως θα μπορούσαν τα άγρια ζώα να «ξέρουν» ποιοι είναι οι συγγενείς τους ; Με άλλα λόγια, ποιους κανόνες συμπεριφοράς ακολουθούν οι οποίοι φανερώνουν έμμεσα ότι ξέρουν την ύπαρξη συγγένειας ;
Ο κανόνας «να είσαι ευγενικός με τους συγγενείς» παρακάμπτει το πραγματικό πρόβλημα, δηλαδή τον τρόπο με τον οποίο θα αναγνωρίζονταν στην πράξη οι συγγενείς. Τα ζώα πρέπει να έχουν πάρει από τα γονίδιά τους έναν απλό κανόνα δράσης τα τελικά αποτελέσματα της οποίας δεν είναι συνειδητά προκαθορισμένα, έναν κανόνα όμως που λειτουργεί τουλάχιστον σε ομαλές συνθήκες. Ολοι οι άνθρωποι ζούμε με κανόνες. Η δύναμή τους είναι τόσο μεγάλη ώστε αν είμαστε στενόμυαλοι ακολουθούμε έναν κανόνα ακόμη κι όταν βλέπουμε καθαρά ότι ούτε εμάς ωφελεί ούτε κανέναν άλλον. Λόγου χάρη, μερικοί φανατικοί Εβραίοι και Μουσουλμάνοι θα προτιμούσαν να λιμοκτονήσουν παρά να παραβούν τον κανόνα που απαγορεύει το χοιρινό κρέας. Ομως ποιους απλούς πρακτικούς κανόνες πρέπει να ακολουθήσουν τα ζώα, οι οποίοι σε κανονικές συνθήκες θα ωφελούσαν έμμεσα τους στενούς συγγενείς τους ;
Αν τα ζώα είχαν την τάση να συμπεριφέρονται αλτρουιστικά σε όλα τα άτομα με τα οποία έχουν φυσική ομοιότητα, θα έκαναν έμμεσα κάποιο καλό στους συγγενείς τους. Τα περισσότερα εξαρτώνται από τις λεπτομέρειες σε κάθε συγκεκριμένη περίπτωση. Εν γένει, ένας τέτοιος κανόνας μόνο στατιστικά θα μπορούσε να οδηγήσει σε «σωστές» αποφάσεις. Αν άλλαζαν οι συνθήκες, αν, λόγου χάρη ένα είδος άρχιζε να ζει σε πολύ μεγαλύτερες ομάδες, αυτός ο κανόνας θα οδηγούσε σε λαθεμένες αποφάσεις. Εννοείται ότι η φυλετική προκατάληψη θα μπορούσε να ερμηνευτεί ως άκριτη γενίκευση μιας τάσης επιλογής συγγενών, να συνταυτίζουμε τον εαυτό μας με άτομα με τα οποία έχουμε φυσική ομοιότητα, και να φερόμαστε άσχημα σε όσα δεν μας μοιάζουν.
Σ’ ένα είδος που τα μέλη του δεν απομακρύνονται πολύ ή απομακρύνονται σε μικρές ομάδες, υπάρχουν πολλές πιθανότητες το τυχαίο άτομο που θα συναντήσετε να είναι στενός συγγενής σας. Σ’ αυτή την περίπτωση ο κανόνας «να είσαι ευγενικός σε κάθε μέλος του είδους σου που θα συναντήσεις» θα είχε θετικό αποτέλεσμα στην επιβίωση, με την έννοια ότι ένα γονίδιο που προδιαθέτει τους υπηκόους του να υπακούουν σ’ αυτόν τον κανόνα θα γινόταν πολυαριθμότερο στη γονιδιακή δεξαμενή. Ισως αυτό εξηγεί την αλτρουιστική συμπεριφορά που αναφέρεται ότι παρουσιάζεται τόσο συχνά σε αγέλες πιθήκων και φαλαινών. Οι φάλαινες και τα δελφίνια πεθαίνουν από ασφυξία αν δεν αναπνεύσουν αέρα. Τα μωρά των φαλαινών ή πληγωμένα άτομα που δεν μπορούν να κολυμπήσουν ως την επιφάνεια βοηθούνται και σώζονται από συντρόφους της αγέλης. Δεν ξέρουμε αν οι φάλαινες έχουν κάποιο τρόπο να ξεχωρίζουν τους στενούς συγγενείς τους, αλλά αυτό ίσως δεν έχει σημασία. Η πιθανότητα δύο τυχαία άτομα της αγέλης να έχουν κάποια συγγένεια μεταξύ τους είναι τόσο μεγάλη ώστε ο αλτρουισμός να αξίζει τον κόπο.
Παρεμπιπτόντως, αναφέρουμε ότι υπάρχει τουλάχιστον μία αυθεντική ιστορία ενός κολυμβητή που κινδύνευε να πνιγεί και τον έσωσε ένα άγριο δελφίνι. Αυτό θα μπορούσε να θεωρηθεί παρανόηση του κανόνα «να σώζεις μέλη της αγέλης». Ο ορισμός ενός μέλους της αγέλης που κινδυνεύει ίσως να είχε κάπως έτσι : «ένα μακρύ πράγμα που σφαδάζει και χτυπιέται στην επιφάνεια του νερού».
Εχει αναφερθεί ότι ενήλικοι αρσενικοί βαβουίνοι υπερασπίζονται με κίνδυνο της ζωής τους την αγέλη εναντίον αρπακτικών, όπως οι λεοπαρδάλεις. Είναι πολύ πιθανό κάθε ενήλικος αρσενικός βαβουίνος να έχει κατά μέσο όρο πολλά κοινά γονίδια με τα άλλα μέλη της αγέλης. Ένα γονίδιο που στην πραγματικότητα «λέει» : «Σώμα, αν είσαι ενήλικο αρσενικό υπερασπίσου την αγέλη από τις λεοπαρδάλεις», θα γινόταν πολυαριθμότερο στη γονιδιακή δεξαμενή. Πριν όμως αφήσουμε αυτό το συχνά αναφερόμενο παράδειγμα, είναι δίκαιο να προσθέσουμε ότι τουλάχιστον μια μεγάλη αυθεντία έχει αναφέρει πολύ διαφορετικά γεγονότα. Σύμφωνα μ’ αυτήν, μόλις εμφανιστεί μια λεοπάρδαλη, οι πρώτοι που εξαφανίζονται από τον ορίζοντα είναι τα ενήλικα αρσενικά.
Τα κλωσσόπουλα μεγαλώνουν σε οικογενειακές ομάδες και ακολουθούν παντού τη μητέρα τους. Βγάζουν δύο είδη φωνών. Εκτός από τα διαπεραστικά τσιρίγματα που ήδη ανέφερα, όταν τρώνε βγάζουν σύντομα μελωδικά τιτιβίσματα. Τα τσιρίγματα με τα οποία επιζητείται η συνδρομή της μητέρας αγνοούνται από τα άλλα κλωσσόπουλα. Τα τιτιβίσματα όμως προσελκύουν και τα άλλα στο μέρος της τροφής : σύμφωνα με το προηγούμενο υποθετικό παράδειγμά μας, τα τιτιβίσματα είναι «κάλεσμα για τροφή». Όπως και εκεί, ο προφανής αλτρουισμός που δείχνουν τα κλωσσόπουλα μπορεί εύκολα να εξηγηθεί ως «επιλογή συγγενών». Επειδή στη φύση όλα τα κλωσσόπουλα μιας κλώσσας είναι κανονικά αδέλφια, το γονίδιο που προκαλεί τα τιτιβίσματα για τροφή θα απλωνόταν, υπό τον όρο ότι το κόστος του κλωσσόπουλου που τιτιβίζει είναι μικρότερο από το μισό του οφέλους που έχουν τα άλλα κλωσσόπουλα. Επειδή το όφελος μοιράζεται σε όλα τα μέλη της ομάδας, που κατά κανόνα είναι περισσότερα από δύο, δεν είναι δύσκολο να δούμε ότι η συγκεκριμένη απαίτηση ικανοποιείται. Φυσικά, ο κανόνας δεν ισχύει σε οικιακές αγροτικές περιπτώσεις, όπου βάζουν την κλώσσα να επωάσει όχι μόνο δικά της αυγά αλλά και πάπιας ή γαλοπούλας. Δεν περιμένουμε βέβαια να το ξέρουν η κλώσσα ή τα κλωσσόπουλα. Η συμπεριφορά τους έχει διαμορφωθεί σύμφωνα με τις συνθήκες που ισχύουν στη φύση, όπου κανονικά δεν υπάρχουν παρείσακτοι στις φωλιές.
Εντούτοις, στη φύση συμβαίνουν περιστασιακά σφάλματα αυτού του τύπου. Στα είδη που ζουν σε κοπάδια ή σε αγέλες, ένα μικρό ορφανό μπορεί να υιοθετηθεί από ένα παράξενο θηλυκό, πιθανότατα από κάποια μητέρα που έχασε το παιδί της. Οσοι ασχολούνται με τους πιθήκους χρησιμοποιούν συχνά τη λέξη «θεία» για το θηλυκό που υιοθετεί κάποιο ορφανό. Στις περισσότερες περιπτώσεις δεν υπάρχει καμία ένδειξη ότι είναι πραγματική θεία ή ότι έχει κάποια άλλη συγγένεια. Αν αυτοί οι ερευνητές ήξεραν για τα γονίδια όσα έπρεπε να ξέρουν, δεν θα χρησιμοποιούσαν τόσο άκριτα τη λέξη «θεία». Οσο συγκινητική κι αν φαίνεται η υιοθεσία, στις περισσότερες περιπτώσεις πρέπει να θεωρείται εκτροπή από έναν θεμελιακό κανόνα, κι αυτό γιατί το γενναιόδωρο θηλυκό όταν φροντίζει το ορφανό δεν ευνοεί τα δικά του γονίδια. Χάνει χρόνο και ενέργεια που θα μπορούσε να τα επενδύσει στη ζωή των συγγενών της, και ειδικότερα των μελλοντικών παιδιών της. Ισως πρόκειται για ένα σφάλμα που συμβαίνει τόσο σπάνια ώστε δεν μπορεί να αλλάξει τους κανόνες κάνοντας το μητρικό ένστικτο περισσότερο επιλεκτικό. Όμως, κατά κανόνα, τέτοιες υιοθεσίες δεν συμβαίνουν και τα ορφανά αφήνονται να πεθάνουν. Υπάρχει ένα τόσο ακραίο παράδειγμα σφάλματος ώστε θα ήταν προτιμότερο να μην το θεωρούμε καθόλου σφάλμα αλλά μαρτυρία εναντίον της θεωρίας του εγωιστικού γονιδίου. Εχουν δει μητέρες πιθηκίνες που είχαν χάσει το μωρό τους να κλέβουν το μωρό κάποιας άλλης και να το φροντίζουν σαν δικό τους. Αυτό το θεωρώ διπλό σφάλμα γιατί η θετή μητέρα όχι μόνο χάνει το χρόνο της αλλά απαλλάσσει ένα ανταγωνιστικό θηλυκό από το βάρος της ανατροφής του παιδιού της, επιτρέποντάς της να αποκτήσει γρήγορα άλλο παιδί. Νομίζω πως πρόκειται για κρίσιμο παράδειγμα που απαιτεί ενδελεχή έρευνα. Πρέπει να μάθουμε πόσο συχνά συμβαίνει κάτι τέτοιο, ποιος είναι ο μέσος δείκτης συγγένειας της θετής μητέρας και του παιδιού, και ποια είναι η στάση της αληθινής μητέρας του παιδιού – σε τελευταία ανάλυση το να υιοθετηθεί το παιδί της είναι προς όφελός της. Μήπως αυτές οι μητέρες προσπαθούν εσκεμμένα να εξαπατήσουν αφελή νεαρά θηλυκά για να υιοθετήσουν τα παιδιά τους ; (Εχει διατυπωθεί η γνώμη ότι οι θετές μητέρες και οι απαγωγείς βρεφών ίσως ωφελούνται, με την έννοια ότι αποκτούν πολύτιμη πείρα στην τέχνη της παιδοκομίας).
Παραδείγματα για τη σκόπιμη άμβλυνση του μητρικού ενστίκτου μας δίνουν οι κούκοι και άλλα πουλιά «παρασιτικά της επώασης», που αφήνουν τα αυγά τους στη φωλιά κάποιου άλλου. Οι κούκοι εκμεταλλεύονται τον κανόνα «να φέρεσαι φιλικά σε κάθε μικρό που βρίσκεται στη φωλιά που έχτισες», ο οποίος είναι εγγενής στους γονιούς-πουλιά. Με εξαίρεση τους κούκους, ο κανόνας έχει πάντα το επιθυμητό αποτέλεσμα να περιορίζει τον αλτρουισμό στους άμεσους συγγενείς, επειδή κατά κανόνα οι φωλιές των πουλιών είναι τόσο απομονωμένες μεταξύ τους ώστε σχεδόν σίγουρα περιέχουν τα μικρά των γονιών. Οι ενήλικοι γλάροι δεν αναγνωρίζουν τα αυγά τους και κάθονται άνετα πάνω στα αυγά άλλων γλάρων ή και σε ψεύτικα αυγά τοποθετημένα από κάποιο φυσιοδίφη. Στη φύση, η αναγνώριση των αυγών δεν είναι πρόβλημα για τους γλάρους επειδή τα αυγά τους δεν μπορεί να κυλήσουν τόσο πολύ ώστε να πλησιάσουν σε κάποια άλλη φωλιά. Εντούτοις, οι γλάροι αναγνωρίζουν τα μικρά τους γιατί τα μικρά, αντίθετα με τα αυγά, μπορεί να απομακρυνθούν και να φτάσουν σε μια γειτονική φωλιά με συχνά μοιραία αποτελέσματα, όπως είδαμε στο πρώτο κεφάλαιο.
Από την άλλη μεριά, οι ουρίες [Εύσωμα στεγανόποδα που ζουν στις ψυχρές χώρες του βόρειου ημισφαίριου. Βαδίζουν δύσκολα στο έδαφος, είναι καλοί κολυμβητές και φωλιάζουν κατά αποικίες στα χείλη απόκρημνων βράχων, κατά μήκος των ακτών (Σ.τ.μ.)] αναγνωρίζουν τα αυγά τους από τα πιτσιλωτά σχέδια που έχουν, και όταν πρόκειται να τα επωάσουν, πετούν τα ξένα. Αυτό κατά πάσα πιθανότητα συμβαίνει γιατί αυτά τα πουλιά κάνουν τις φωλιές τους σε ομαλούς βράχους και υπάρχει κίνδυνος να κυλήσουν και να μπερδευτούν με άλλα. Θα μπορούσε λοιπόν να ρωτήσει κανείς : γιατί μπαίνουν στο κόπο να ξεχωρίζουν τα ξένα και να κάθονται μόνο στα δικά τους ; Προφανώς θα ήταν δυνατό κάθε θηλυκό να επωάζει οποιοδήποτε αυγό, οπότε δεν θα υπήρχε θέμα αν είναι δικό της ή ξένο. Κι αυτό ακριβώς υποστηρίζουν οι οπαδοί της επιλογής ομάδων. Αλλά ας εξετάσουμε τι θα συνέβαινε αν εξελισσόταν ένας τέτοιος κύκλος ομαδικής επώασης. Ο μέσος αριθμός παιδιών για τις ουρίες είναι ένα. Αυτό σημαίνει ότι για να δουλέψει αποτελεσματικά ο κύκλος της αμοιβαίας επώασης, κάθε ενήλικο θηλυκό πρέπει να επωάσει κατά μέσο όρο ένα αυγό. Ας υποθέσουμε πως κάποια έκανε την κατεργαριά και δεν καθόταν σε ένα αυγό. Αντί να χάνει χρόνο στην επώαση ξόδευε τον καιρό της να γεννά περισσότερα αυγά και για μεγάλη της χαρά άλλα περισσότερο αλτρουιστικά θηλυκά τα φρόντιζαν για λογαριασμό της. Αυτά τα θηλυκά θα συνέχιζαν να υπακούουν πιστά στον κανόνα : «Αν δεις κάποιο παρατημένο αυγό κοντά στη φωλιά σου, να το μαζέψεις και να το επωάσεις». Σ’ αυτή την περίπτωση το γονίδιο για την κατεργαριά θα απλωνόταν στον πληθυσμό, οπότε θα έσπαγε ο όμορφος κύκλος της φιλικής επώασης.
Θα μπορούσε όμως κάποιος να διερωτηθεί τι θα γινόταν αν τα έντιμα θηλυκά ανταπέδιδαν και δεν υπέκυπταν στον εκβιασμό επωάζοντας ένα και μόνο αυγό ; Αυτό θα ανέτρεπε τα σχέδια των «κατεργάρικων» γιατί θα έβλεπαν τα αυγά τους παρατημένα στους βράχους να μην τα επωάζει κανείς. Μήπως αυτό θα τα επανέφερε σε τάξη ; Δυστυχώς όχι. Εφόσον δεχτήκαμε ότι τα θηλυκά δεν ξεχωρίζουν το ένα αυγό από το άλλο, αν τα έντιμα πουλιά εφάρμοζαν την παραπάνω τακτική, πιθανώς και τα δικά τους αυγά να είχαν την ίδια τύχη με αυτά των κατεργάρικων. Ομως τα κατεργάρικα θα εξακολουθούσαν να πλεονεκτούν. Θα γεννούσαν περισσότερα αυγά και θα επιβίωναν περισσότερα μικρά τους. Τα έντιμα θηλυκά για να εξουδετερώσουν τα κατεργάρικα δεν έχουν άλλο τρόπο από το να ξεχωρίζουν με σιγουριά τα αυγά τους. Με άλλα λόγια, να πάψουν να είναι αλτρουιστικά και να ενδιαφέρονται μόνο για το δικό τους συμφέρον.
Για να χρησιμοποιήσω τη γλώσσα του Maynard Smith, η αλτρουιστική στρατηγική της υιοθεσίας δεν είναι εξελικτικά σταθερή. Είναι ασταθής, με την έννοια ότι μπορεί να βελτιωθεί με την εγωιστική αντιστρατηγική το θηλυκό να γεννά περισσότερα αυγά απ’ όσα πρέπει και να μην τα επωάζει. Αλλά και αυτή η εγωιστική αντιστρατηγική είναι με τη σειρά της ασταθής, γιατί η αλτρουιστική στρατηγική στην οποία στηρίζεται είναι ασταθής, συνεπώς θα εξαφανιστεί και αυτή. Η μόνη εξελικτικά σταθερή στρατηγική για την ουρία είναι να αναγνωρίζει το αυγό της και να επωάζει αποκλειστικά το δικό της. Κι αυτό ακριβώς συμβαίνει.
Τα είδη των ωδικών πτηνών που εκμεταλλεύονται οι κούκοι αντέδρασαν : δεν έμαθαν να αναγνωρίζουν τα αυγά τους και να ευνοούν όσα αυγά έχουν ειδικά στίγματα. Αυτή η στρατηγική είναι αποτελεσματική γιατί τα θηλυκά δεν διατρέχουν τον κίνδυνο να τα εκμεταλλευτούν άλλα άτομα του είδους τους. Αλλά και οι κούκοι αντεπιτέθηκαν με τη σειρά τους, κάνοντας αυγά που μοιάζουν όλο και περισσότερο ως προς το μέγεθος, το χρώμα και τα στίγματα με εκείνα των ξενιστών τους. Αυτό αποτελεί παράδειγμα μιας απάτης που συχνά αποδίδει. Το αποτέλεσμα αυτού του συγκεκριμένου εξελικτικού ανταγωνισμού ήταν η αξιοσημείωτη τελειότητα μίμησης που παρατηρείται στα αυγά του κούκου. Μπορούμε να υποθέσουμε ότι ένας αριθμός αυγών και νεοσσών του κούκου «ανακαλύπτεται». Οσα όμως δεν ανακαλύπτονται, επιζούν και γεννούν την επόμενη γενεά. Μ’ αυτόν τον τρόπο στο γονιδιακό απόθεμα του κούκου εξαπλώνονται τα γονίδια για αποτελεσματικότερη εξαπάτηση. Με τον ίδιο τρόπο, και τα πουλιά-ξενιστές, που έχουν αρκετή παρατηρητικότητα ώστε να διακρίνουν την παραμικρή ατέλεια στην απομίμηση των αυγών τους, είναι αυτά που συνεισφέρουν τα περισσότερα στο δικό τους γονιδιακό απόθεμα. Ετσι, τα παρατηρητικά και δύσπιστα μάτια περνούν στην επόμενη γενεά τους. Αυτό είναι ένα καλό παράδειγμα του τρόπου με τον οποίο η φυσική επιλογή οξύνει την αντίληψη λεπτών διαφορών : σ’ αυτή την περίπτωση, των διαφορών εναντίον ειδών που τα μέλη τους κάνουν τα πάντα για να τις εξαφανίσουν.
Ας επιστρέψουμε τώρα στην σύγκριση της «εκτίμησης» που κάνει ένα ζώο για τη συγγένειά του με άλλα μέλη της ομάδας του και της αντίστοιχης εκτίμησης που κάνει ένας έμπειρος ερευνητής φυσιοδίφης. Ο Brian Bertram ξόδεψε πολλά χρόνια μελετώντας στο εθνικό πάρκο του Serengeti [Εκταση 13.000 τ.χλμ. στο βόρειο τμήμα της Τανζανίας και νοτιοανατολικά της Λίμνης Βικτώριας (Σ.τ.μ.)] τη βιολογία των λιονταριών. Στηριζόμενος στις γνώσεις του για τις αναπαραγωγικές συνήθειές τους, εκτίμησε τον μέσο δείκτη συγγένειας ανάμεσα στα άτομα μιας τυπικής αγέλης. Τα γεγονότα στα οποία στηρίχθηκε για να κάνει τις εκτιμήσεις του είναι τα εξής : Μια τυπική αγέλη λιονταριών περιλαμβάνει περίπου 7 ενήλικα θηλυκά, που είναι και τα μονιμότερα μέλη της, και 2 αδέσποτα αρσενικά. Περίπου τα μισά ενήλικα θηλυκά γεννούν σχεδόν ταυτόχρονα και μεγαλώνουν από κοινού τα μικρά τους, έτσι ώστε είναι δύσκολο να ξέρει κανείς τη μητέρα κάθε μικρού. Ο τυπικός αριθμός νεογνών σε κάθε γέννα είναι 3. Η πατρότητα των μικρών μοιράζεται εξίσου στα ενήλικα αρσενικά της αγέλης. Τα νεαρά θηλυκά παραμένουν στην αγέλη και αντικαθιστούν τα γέρικα θηλυκά που πεθαίνουν ή που φεύγουν. Τα νεαρά αρσενικά εγκαταλείπουν την ομάδα όταν φτάσουν στην εφηβεία. Όταν είναι μεγάλα περιπλανώνται από ομάδα σε ομάδα κατά μικρές συγγενικές «συμμορίες» ή σε ζεύγη, και είναι απίθανο να επιστρέψουν στην αρχική οικογένεια.
Με τις παραπάνω και άλλες παραδοχές είναι δυνατό να βρεθεί μια μέση τιμή του δείκτη συγγένειας δύο ατόμων μιας τυπικής αγέλης λιονταριών. Ο Bertram για ένα τυχαίο ζευγάρι αρσενικών βρίσκει τη τιμή 0,22 και για ένα ζευγάρι θηλυκών 0,15. Είναι σαν να λέμε ότι τα αρσενικά ενός κοπαδιού κατά μέσο όρο είναι κάτι λιγότερο από ετεροθαλή αδέλφια, και τα θηλυκά κάτι περισσότερο από πρωτοξάδελφα. Φυσικά, είναι δυνατό κάποιο συγκεκριμένο ζευγάρι ατόμων να είναι πραγματικά αδέλφια αλλά ο Bertram δεν μπορούσε να το ξέρει, και είναι σίγουρο πως και τα λιοντάρια δεν το ξέρουν. Από την άλλη μεριά, και οι μέσες τιμές συγγένειάς τους που βρήκε είναι κατά κάποιο τρόπο «γνωστές» και στα λιοντάρια. Αν αυτοί οι αριθμοί είναι αντιπροσωπευτικοί για μια μέση αγέλη, τότε κάθε γονίδιο που θα προδιέθετε τα αρσενικά να συμπεριφέρονται στα άλλα αρσενικά στα άλλα αρσενικά σαν να ήταν σχεδόν ετεροθαλή αδέλφια, θα είχε θετική αξία για την επιβίωση. Κάθε γονίδιο που θα προχωρούσε περισσότερο και θα έκανε τα αρσενικά να συμπεριφέρονται με φιλικότερο τρόπο απ’ ό, τι στα κανονικά αδέλφια, κατά μέσο όρο θα τιμωρούνταν όπως ακριβώς κι αν δεν φερόταν αρκετά φιλικά (αν, λόγου χάρη, συμπεριφερόταν στα άλλα αρσενικά σαν να ήταν δευτεροξάδελφα). Αν τα γεγονότα στη ζωή των λιονταριών έχουν όπως λέει ο Bertram, και αν είχαν έτσι για πολλές γενεές, τότε μπορούμε να περιμένουμε ότι η φυσική επιλογή θα είχε ευνοήσει ένα βαθμό αλτρουισμού σύμφωνα με τη μέση τιμή του δείκτη συγγένειας μιας τυπικής αγέλης. Αυτό ακριβώς εννοούσα όταν έλεγα ότι οι εκτιμήσεις για τις συγγένειες που κάνουν ένα ζώο και ένας καλός ερευνητής φυσιοδίφης πρέπει τελικά να είναι οι ίδιες.
Ετσι φτάνουμε στο συμπέρασμα ότι στην εξέλιξη του αλτρουισμού η «πραγματική» συγγένεια μπορεί να είναι λιγότερο σημαντική από μια καλή εκτίμηση που κάνει ένα ζώο για τη συγγένεια. Αυτό αποτελεί πιθανώς το κλειδί για να καταλάβουμε γιατί στη φύση η φροντίδα των γονιών για τα παιδιά τους είναι πολύ περισσότερο διαδεδομένη και πολύ εντονότερη από τον αλτρουισμό αδελφού/αδελφής, και γιατί τα ζώα δίνουν πολύ μεγαλύτερη αξία στους εαυτούς τους απ’ όση στα αδέλφια τους. Εν συντομία, θέλω να τονίσω ότι εκτός από το δείκτη συγγένειας πρέπει να εξετάζουμε και κάτι που θα το λέγαμε δείκτη «βεβαιότητας». Μολονότι από γενετική άποψη ησχέση γονιού/παιδιού δεν είναι στενότερη από τη σχέση αδελφού/αδελφής, η «βεβαιότητά» της είναι μεγαλύτερη. Κανονικά, είστε πολύ πιο σίγουροι ποια είναι παιδιά σας παρά ποια είναι αδέλφια σας. Και ακόμη πιο σίγουροι ότι είστε ο εαυτός σας !
Μιλήσαμε για κατεργάρικα θαλασσοπούλια και στα επόμενα κεφάλαια θα πούμε περισσότερα για ψεύτες, κατεργάρηδες και εκμεταλλευτές. Σ’ έναν κόσμο όπου πάντα βρίσκονται άτομα έτοιμα να εκμεταλλευτούν τις ευκαιρίες που προσφέρει ο μεταξύ συγγενών αλτρουισμός και να τον χρησιμοποιήσουν προς όφελός τους, μια μηχανή επιβίωσης πρέπει να εξετάζει ποιον μπορεί να εμπιστεύεται, για ποιον μπορεί να είναι σίγουρη. Αν ο Β είναι πραγματικά ένα μικρότερο αδελφάκι μου, τότε πρέπει να νοιάζομαι γι’ αυτό το μισό απ’ όσο για τον εαυτό μου και ακριβώς όσο για το παιδί μου. Όμως μπορώ να είμαι τόσο σίγουρος γι’ αυτό όσο για το παιδί μου ; Πως μπορώ να ξέρω ότι είναι αδελφάκι μου ;
Αν ο Δ είναι αυθεντικός δίδυμος αδελφός μου, θα έπρεπε να νοιάζομαι γι’ αυτόν διπλάσια απ’ όσο για το παιδί μου, δηλαδή να θεωρώ τη ζωή του το ίδιο πολύτιμη με τη δική μου. Όμως μπορώ να είμαι σίγουρος γι’ αυτόν ; Είναι βέβαιο πως μου μοιάζει αλλά αυτό μπορεί να οφείλεται στο ότι έχουμε κοινά μόνο όσα γονίδια αφορούν τα χαρακτηριστικά του προσώπου. Οχι, δεν θα θυσιάσω τη ζωή μου για χάρη του γιατί αν υπάρχει μόνο κάποια πιθανότητα να έχει το 100% των γονιδίων μου, εγώ είμαι απόλυτα σίγουρος πως έχω 100% των γονιδίων μου, οπότε αξίζω περισσότερο από αυτόν. Είμαι το μόνο άτομο για το οποίο είναι σίγουρο το καθένα από τα εγωιστικά μου γονίδια. Μολονότι θεωρητικά ένα γονίδιο για ατομικό αλτρουισμό θα μπορούσε να εκτοπιστεί από κάποιο ανταγωνιστικό γονίδιο «για αλτρουιστική διάσωση» τουλάχιστον ενός αυθεντικού διδύμου, δύο παιδιών ή αδελφών ή τουλάχιστον 4 εγγονιών κλπ., το γονίδιο του ατομικού εγωισμού έχει το τεράστιο πλεονέκτημα ότι είναι βέβαιο για την ταυτότητα του ατόμου. Το ανταγωνιστικό γονίδιο για τον αλτρουισμό προς τους συγγενείς διατρέχει τον κίνδυνο να κάνει λάθη ως προς την ταυτότητα των ατόμων είτε πραγματικά τυχαία είτε γιατί τα προκάλεσαν σκόπιμα κατεργάρηδες και παράσιτα. Συνεπώς, στη φύση πρέπει να περιμένουμε τον ατομικό εγωισμό σε βαθμό μεγαλύτερο από αυτόν που θα προβλέπαμε αν υπολογίζαμε μόνο τη γενετική συγγένεια.
Σε πολλά είδη του ζωικού βασιλείου η μητέρα είναι περισσότερο σίγουρη από τον πατέρα ότι τα παιδιά είναι δικά της. Η μητέρα γεννά ορατά, χειροπιαστά αυγά ή κουβαλά μέσα της το παιδί. Εχει μεγάλη πιθανότητα να ξέρει τους φορείς των γονιδίων της. Ο ατυχής πατέρας μπορεί να απατηθεί πολύ εύκολα. Πρέπει λοιπόν να περιμένουμε οι πατέρες να καταβάλλουν μικρότερη προσπάθεια από τις μητέρες για τη φροντίδα των παιδιών. Στο ένατο κεφάλαιο, που ασχολείται με τον ανταγωνισμό των φύλων, θα δούμε ότι συντρέχουν και άλλοι λόγοι που οδηγούν στο ίδιο συμπέρασμα. Για τον ίδιο λόγο οι «εκ μητρός» γιαγιάδες είναι πιο σίγουρες για τα εγγόνια τους συγκριτικά με τις «εκ πατρός» γιαγιάδες και πρέπει να περιμένουμε να δείχνουν μεγαλύτερο αλτρουισμό. Κι αυτό γιατί οι εκ μητρός γιαγιάδες είναι σίγουρες ότι τα παιδιά των θυγατέρων τους είναι δικά τους, ενώ για τις εκ πατρός γιαγιάδες υπάρχει το ενδεχόμενο να απατήθηκε ο γιος τους.
Οι εκ μητρός παππούδες έχουν τόση σιγουριά για τα εγγόνια τους όση και οι εκ πατρός γιαγιάδες, επειδή και οι δύο υπολογίζουν μια γενεά βεβαιότητας και μια αβεβαιότητας. Παρομοίως, οι θείοι από τη μεριά της μητέρας θα έπρεπε να ενδιαφέρονται περισσότερο για τους ανιψιούς και τις ανιψιές τους συγκριτικά με τους θείους από τη μεριά του πατέρα, και γενικά να δείχνουν τον ίδιο αλτρουισμό με τις θείες. Πραγματικά, σε μια κοινωνία με υψηλό δείκτη συζυγικής απιστίας, οι εκ μητρός θείοι έπρεπε να είναι περισσότερο αλτρουιστές από τους «πατέρες», επειδή έχουν περισσότερα ερείσματα για τη συγγένειά τους με το παιδί [Η παρατήρηση προστέθηκε το 1978. Παρέβλεψα το γεγονός ότι ο R.D. Alexander είχε αναφερθεί στο θέμα το 1974]. Ξέρουν ότι η μητέρα του παιδιού είναι τουλάχιστον ετεροθαλής αδελφή τους. Ο «νόμιμος» πατέρας δεν ξέρει τίποτε. Δεν γνωρίζω αν υπάρχουν στοιχεία που να στηρίζουν αυτές τις προβλέψεις, τις δίνω όμως με την ελπίδα ότι άλλοι θα ψάξουν ή θα αρχίσουν να ψάχνουν να τα βρουν.
Ειδικότερα, οι ανθρωποκοινωνιολόγοι ίσως μας έλεγαν ενδιαφέροντα πράγματα. Το γεγονός ότι ο αλτρουισμός των γονέων είναι συνηθέστερος από τον αδελφικό αλτρουισμό εξηγείται λογικά με βάση το «πρόβλημα της ταυτότητας». Εντούτοις, με αυτό δεν εξηγείται η θεμελιώδης ασυμμετρία που υπάρχει στη σχέση ανάμεσα στον πατέρα και το παιδί. Οι γονείς φροντίζουν περισσότερο τα παιδιά τους απ’ όσο τα παιδιά τους γονείς τους, μολονότι η γενετική σχέση είναι συμμετρική και η βεβαιότητα συγγένειας ίδια και προς τις δύο κατευθύνσεις. Ενας λόγος αυτής της ασυμμετρίας είναι ότι οι γονείς, από πρακτική άποψη, βρίσκονται σε καλύτερη θέση να βοηθήσουν τα παιδιά τους, γιατί είναι μεγαλύτεροι και πιο έμπειροι στα προβλήματα της ζωής. Ακόμη κι αν ήθελε ένα μωρό να θρέψει τους γονείς του, δεν έχει τα κατάλληλα εφόδια να το κάνει.
Στη σχέση πατέρα/παιδιού υπάρχει μια ακόμη ασυμμετρία που δεν απαντάται στη σχέση αδελφού/αδελφής. Τα παιδιά έχουν πάντα μικρότερη ηλικία από τους γονείς τους. Αυτό σημαίνει ότι συχνά, όχι πάντα βέβαια, περιμένουν να ζήσουν περισσότερο. Όπως τόνισα παραπάνω, η προσδοκία ζωής είναι σημαντικός παράγοντας ο οποίος, και στον καλύτερο από τους δυνατούς κόσμους υπεισέρχεται στον «υπολογισμό» που κάνει ένα ζώο όταν «αποφασίζει» αν θα συμπεριφερθεί αλτρουιστικά ή εγωιστικά. Σ’ ένα είδος στο οποίο τα παιδιά έχουν μεγαλύτερη μέση διάρκεια ζωής από τους γονείς, κάθε γονίδιο για αλτρουιστική συμπεριφορά του παιδιού προς τον πατέρα θα λειτουργούσε μειονεκτικά. Θα ήταν μια αφύσικη αλτρουιστική αυτοθυσία προς όφελος ατόμων που βρίσκονται πλησιέστερα στο θάνατο από γηρατειά απ’ όσο ο αλτρουιστής. Από την άλλη μεριά, ένα γονίδιο για αλτρουιστική συμπεριφορά του πατέρα προς το παιδί θα λειτουργούσε πλεονεκτικά στο πλαίσιο των προσδοκιών ζωής.
Μερικοί εκφράζουν τη γνώμη ότι η επιλογή συγγενών είναι πολύ καλή ως θεωρία αλλά στην πράξη λίγα μόνο παραδείγματα δείχνουν ότι λειτουργεί. Όμως αυτή η κριτική γίνεται μόνο απ’ όσους δεν κατανοούν την πραγματική σημασία της επιλογής συγγενών. Η αλήθεια είναι ότι όλα τα παραδείγματα προστασίας του παιδιού και γονικής φροντίδας, και όλα τα σχετικά σωματικά όργανα, π.χ. γαλακτογόνοι αδένες, μάρσιποι των καγκουρώ κ.ά., είναι παραδείγματα λειτουργίας στη φύση της αρχής της επιλογής συγγενών. Βέβαια, οι επικριτές είναι εξοικειωμένοι με την απανταχού παρατηρούμενη γονική φροντίδα αλλά δεν καταλαβαίνουν ότι, όπως και ο αλτρουισμός αδελφού/αδελφής, είναι και αυτή παράδειγμα επιλογής συγγενών. Όταν λένε ότι θέλουν παραδείγματα, εννοούν ότι θέλουν παραδείγματα διαφορετικά από τη γονική φροντίδα, και είναι αλήθεια πως αυτά είναι λιγότερο συνηθισμένα. Εχω υποδείξει μερικές αιτίες αυτής της κατάστασης. Όμως θα ξεστράτιζα αν ανέφερα παραδείγματα αλτρουισμού μεταξύ αδελφού/αδελφής – πραγματικά υπάρχουν λίγα. Εντούτοις, δεν θα το κάνω γιατί θα ενίσχυα τη λαθεμένη ιδέα (προσφιλή στον Wilson, όπως είδαμε) ότι η επιλογή συγγενών εφαρμόζεται ειδικά σε σχέσεις διαφορετικές από αυτή του πατέρα/παιδιού.
Οι λόγοι που οδήγησαν σ’ αυτό το σφάλμα είναι κυρίως ιστορικοί. Το εξελικτικό πλεονέκτημα της γονικής φροντίδας είναι τόσο προφανές ώστε δεν χρειαζόταν να περιμένουμε τον Hamilton να το επισημάνει. Είχε κατανοηθεί ήδη από την εποχή του Δαρβίνου. Όταν ο Hamilton αποδείκνυε τη γενετική σημασία τους, ήταν φυσικό να δίνει βαρύτητα ακριβώς στις άλλες σχέσεις. Ειδικότερα, πήρε παραδείγματα από τη ζωή κοινωνικών εντόμων, όπως τα μυρμήγκια και οι μέλισσες, στα οποία, καθώς θα δούμε σε επόμενο κεφάλαιο, η σχέση αδελφής/αδελφής είναι ιδιαίτερα σημαντική. Άκουσα μάλιστα μερικούς να λένε ότι η θεωρία του Hamilton εφαρμόζεται μόνο στα κοινωνικά έντομα !
Αν κάποιος αρνείται να παραδεχτεί ότι η γονική φροντίδα είναι ενεργό παράδειγμα επιλογής συγγενών, είναι ενεργό παράδειγμα επιλογής συγγενών, είναι υποχρεωμένος να διατυπώσει μια γενική θεωρία φυσικής επιλογής η οποία να προβλέπει τον αλτρουισμό των γονιών στα παιδιά τους αλλά να μη προβλέπει αλτρουιστική συμπεριφορά μεταξύ πλάγιων συγγενών. Πιστεύω ότι θα αποτύχει.
Richard Dawkins – ΤΟ ΕΓΩΙΣΤΙΚΟ ΓΟΝΙΔΙΟ
Αντικλείδι , https://antikleidi.com
Συναφές:

